Animal Communication (not limited to birds)
"Linguistic analogies and behavior: the finite-state behavioral grammar of food-hoarding in hamsters", by C. H. Jones, J. P. J. Pinel, Berhavioural Brain Research, Jan. 22 1990, 36, 3, 189 - 197
The authors use the same methods of linguistic analysis (finite state grammar) to study the food-hoarding behavior of Syrian golden hamsters (Mesocricetus auratus). The authors explicitly note that "...most behavioral research has focused not on behavioral sequences themselves, but on a few isolated measures of the behavior or its outcomes...". The authors follow the methodology of Rodger and Rosebrugh and other scientists (see "Emergent Computation: Emphasizing Bioinformatics", pages 348-357). The authors chose the food-hoarding behavior of hamsters as the motor aspects of this behavior could be broken down into sequences of identifiable behavioral motor elements that could be observed, and because previous observation of hamsters that this behavior of hamsters is affected by brain lesions. The beginning of a hoarding sequence is defined as the point at which a hamster inserted food into its cheek pouches in the foraging area, until the food was ejected in the home compartment. Details about the experiment are not discussed here, but the results follow.
Terminal Vocabulary for Syrian Golden Hamster Food-hoarding

Observed Type-3 Statements of Syrian Golden Hamster Food-hoarding Behavior

Hamster Non-Deterministic Finite State Machine

The authors identify what they feel are linguistic "language-reducing" phrases. Linguisic analysis led the authors to two hypotheses:
- Hamsters often explore the foraging area after filling their cheek pouches with food, either to assess the amount of remaining food or to identify potential hazards.
- Hamsters that have carried food back to their nest area often inspect several potential deposition sites before emptying their cheek pouches.
DFSA for Efficient Foraging
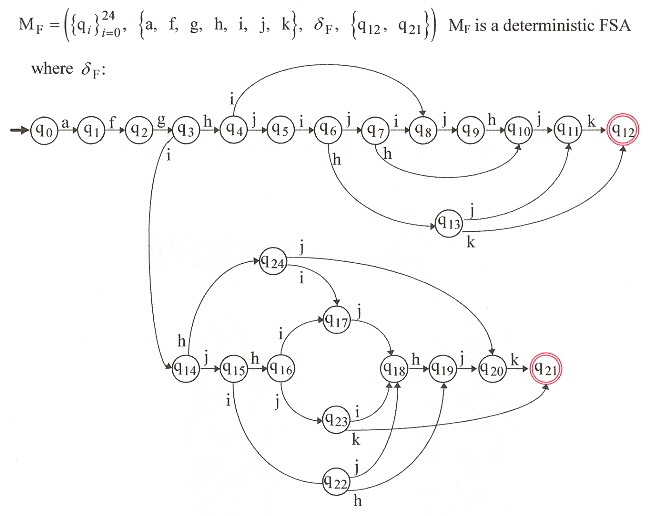
Type-3 Grammar for Efficient Foraging

DFSA and Type-3 Grammar for Efficiency at Home
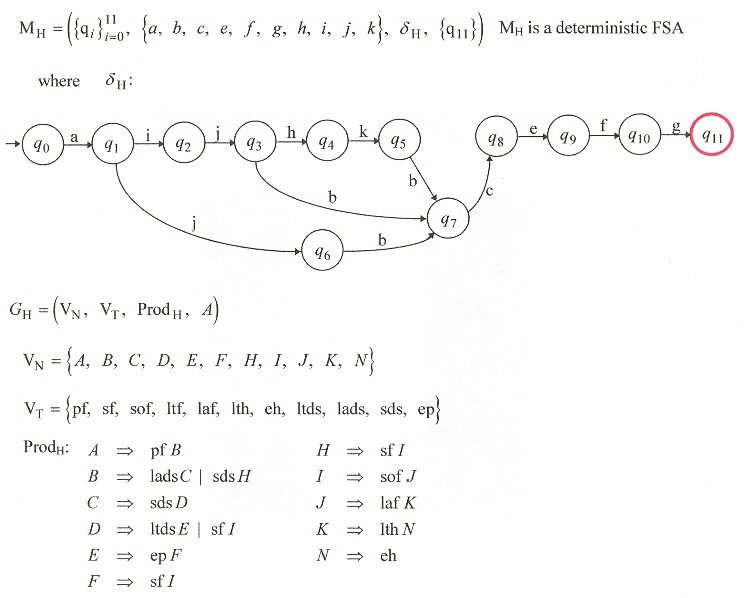
DFSA and Type-3 Grammar for Inefficiency both for Foraging and at Home
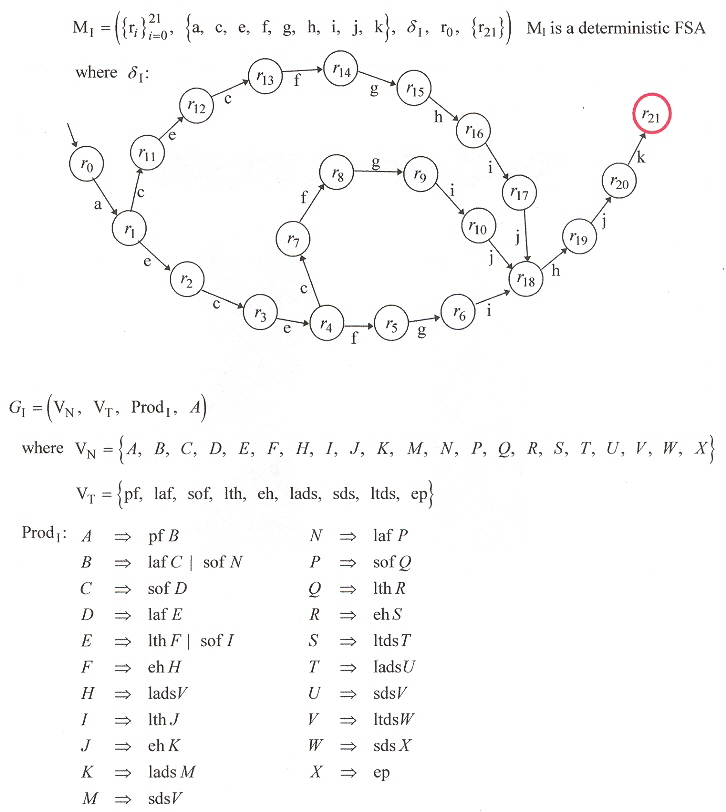
Thus in Figure 4,
1/ PF→ 2/ LTH–EH→ 13/ LADS–LTDS–SDS–EP→ 0/
While this corresponds to sequence "a, f, g, i, h, j, k" or in GF, this is corresponds to:
S ⇒ pf A ⇒
pf · lth B ⇒
pf · lth · eh C ⇒
pf · lth · eh · lads Q ⇒
pf · lth · eh · lads · ltds O ⇒
pf · lth · eh · lads · ltds · sds Y
⇒ pf · lth · eh · lads · ltds · sds · ep
Stridulation is the creation of sound by rubbing body parts together. Stridulation is a fairly common method of producing sound that can be used for communication for various purposes such as mating, defense, territorial claims, distraction to protect young, etc. Stridulation as a means of communication might possibly be found to have enough structure to constitute a language or to evolve to such a level of complexity. Stridulation has been found in tenrecs, a mammal a; black fire ants Solenopsis richterib, earth-boring dung beetles Geotrupidae c; stag beetles Cleoptera, Lucanu cervus d; crickets Gryllidae e; grasshoppers Gomphocerinae f; cicadas Hemiptera, Homoptera g; channel catfish Siluroidei: Clariidae h; club-winged Manakains Machaeropterus deliciosus i; etc.
a "Orders and Families of Recent Mammals of the World", by S. Anderson, J. K. Jones Jr., John Wiley and Sons, 1984, 117 - 144
b "Analysis of acoustic communication by ants", by R. Hickling, R. L. Brown, Journal of the Acoustic Society of America, 2000, 108, 4, 1920 - 1929
c "Dung beetles (Coleoptera: Scarabaeidae), monkeys, and dung beetle conservation in amazonia", by K. Vulinec, Florida Entomology, 2000, 83, 229 - 241
d "Untersuchungen zum Stridulationsverhalten der Hirschkäfer-Larven (Lucanus cervus L.) (Coleoptera: Lucanidae)", by E. Sprecher-Uebersax, Mitteilungen der Schweizerischen Entomologischen Gesellschaft, 1998, 71, 471 - 479
e "Stridulation and tegminal resonance in the tree cricket Oecanthus nigricornis (Orthoptera: Gryllidae: Oeceanthinae)", by E. Sismondo, Comparative Physiology, 1979, A 129, 269 - 279
f "Pharmacological Activation of Stridulation in Grasshopper Chorthippus albomargginatus (Orthoptera: Gomphocerinae)", by V. Y. Vedenina, R. Heinrich, N. Elsner, Journal of Evolutionary Biochemistry and Physiology, July 2001, 37, 4, 388 - 397
g "Analysis of Acoustic Signals Produced by the Cicada Platypedia putnami Variety lutea (Homoptera: Tibicinidae)", by A. F. Sanborn, P. K. Phillips, Annals of the Entomological Society of America, May 1999, 92, 3, 451 - 455
h "The evolutionary origin and functional divergence of sound production in fish: catfish stridulation mechanisms", by I. Kaatz, Journal of Morphology, 1997, 232(3): 272
i "Courting Bird Sings With Stridulating Wing Feathers", by K. S. Bostwick, R. O. Prum, Science, July 29 2005, 309, 736
"Sensory Systems and Spatial Memory in the Fruit Bat Rousettus aegyptiacus", by R. A. Holland, P. Winter, D. A. Waters, Ethology, August 2005, 111, 8, 715 - 725
The megachiroptteran fruit bat Rousettus aegyptiacus is able to orient and navigate using both vision and echolocation. However, echolocation works at comparatively short range when compared to vision. It is also mentioned in this paper that there is evidence that dolphins (cetaceans) also utilize both vision and echolocation. It should be noted that dolphins have been trained to utilize complicated human command languages.
"Language Evolution: What do Chimpanzees Have to Say", by A. C. Arcadi, Current Biology, 2005, 15, 21, R884 - R886
The author limits languages animals, to possible non-humans, however. After all, dealing with the evolution of languages does require a extension of languages beyond humans. The author points out that examples of non-human languages (even if the languages are limited), have been found in a variety of non-humans, as follows.
- Vervet monkeys (Cercopithecus aethiops) a, b
- Ring-tailed lemurs (Lemur catta) c
- Campbell's monkeys (C. diana and C. campbelli) d
- Domestic chickens (Gallus gallus) e
- Chimpanzees f
The author points out that vervet monkeys have three distinct alarm calls that identify three different predators (eagles, leopards, snakes).
While the author points out that such languages (he limits his analysis only to "words", ignoring grammatical structures found. For example, in birds, languages with grammatical structures do appear to be what has been found g). The author points out that, indeed, chimpazees appear to have multiple, different "food calls" (the 'rough grunt'). The 'rough grunt' depends upon amount and preference of the food type.
a "Auditory communication among vervet monkeys (Cercopithecus aethiops)", by T. T. Struhsaker, 1968, in "Social Communication among Primates", Ed. by S. A. Altmann, Univ. of Chicago Press, 281 - 324
b "Monkey responses to three different alarm calls: Evidence of predator classification and semantic communication", by R. M. Seyfarth, D. L. Cheney, P. Marler, Science, 1980, 210, 801 - 803
c "What is communicated in the antipredator calls of lemurs: Evidence from playback experiments with ring-tailed lemurs and ruffed lemurs", by J. Macedonia, Ethology, 1990, 86, 177 - 190
d "Referential signaling in non-human primates: Cognitive precursors and limitations for the evolution of language", by Zuberbrühler, Advanced Studies of Behavior, 2003, 33, 265 - 307
e "On the meaning of alarm calls: Functional reference in an avian vocal system", by C. S. Evans, L. Evans, P. Marler, Animal Behavior, 1993, 46, 23 - 38
f "Functionally referential communication in a chimpanzee", by S. Slocombe, K. Zuberbrühler, Current Biology, 2005, 15, 1779 - 1784
g "A phonological analysis of male gibbon singing behavior", by J. C. Mitani, P. Marler, Behaviour, 1989, 109, 20 - 45
"Ultrviolet signals fighting ability in a lizard", by J. Stapley, M. J. Whiting, Biology Letters, Feb. 28 2006, 1466 (DOI:10.1098/rsbl.2005.0419)
Ultraviolet light has been shown to be of significamce in the territorial communication behaviour of Pomacentridae (damselfish), and mate choice with various birds (pied flycatcher, Parus caeruleus). Fighting behaviour has also been found in lizards a. The throat of the male Platysaurus broadleyi (African flat lizard) is highly reflective in the ultraviolet range. Throat displays affect fighting (for territory or mates, for example), thus ultraviolet reflectance constitutes a not uncommon form of form of animal communication.
a "Ultrviolet signals fighting ability in a lizard", New York Times, Science Section, Jan. 10, 2006.
"Thermal Modeling of Snake Infrared Reception: Evidence for a Limited Detection Range", by B. S. Jones, W. F. Lynn, M. O. Stone, Journal of Theoretical Biology, 2001, 209, 201 - 211
What are some of the parameters found in the sensitivity and sensing range of the facial pit organs of Boidae (pythons) and Crotalinae (pit vipers)? Pit organs are facial pits lined with infrared-sensing receptor cells. Pit organs are utilized for the detection of prey, dodging predators, and finding basking spots. Pit organ sensitivity has been determined to be 0.003° C, with a response time of 0.05° C/sec. Detection range varies roughly between 5 cm. to 50 cm. (depending upon species). Thus heat detection via infrared radiation works much like sight and could take part in a system of biological communication.
"Smell in Sterio? Most of Us Would Just Say No", by J. Gorman, New York Times Science Section, 2/7/2006, p. D4 (Science, Raghav Rajan, James P. Clement, Upinder S. Bhalla, University of Agricultural Science, Bangalore)
Using brain probes, researchers found that in rats, the sensory information from each nostril was processed separately in the brain, giving rats enough data to determine location using only their sense of smell.
Back to TopInsect Communication
"Neural mechanisms in insect navigation: polarization compass and odometer", Labhart T, Meyer EP., Current Opinion in Neurobiology, 2002 Dec;12(6):707-14, Zoologisches Institut der Universitat, Winterthurerstr. 190, CH-8057 Zurich, Switzerland. labhart@zoo.unizh.ch
Insect navigation relies on path integration, a procedure by which information about compass bearings pursued and distances travelled are combined to calculate position. Three neural levels of the polarization compass, which uses the polarization of skylight as a reference, have been analyzed in orthopteran insects. A group of dorsally directed, highly specialized ommatidia serve as polarization sensors. Polarization-opponent neurons in the optic lobe condition the polarization signal by removing unreliable and irrelevant components of the celestial stimulus. Neurons found in the central complex of the brain possibly represent elements of the compass output. The odometer for measuring travelling distances in honeybees relies on optic flow experienced during flight, whereas desert ants most probably use proprioreceptive cues.
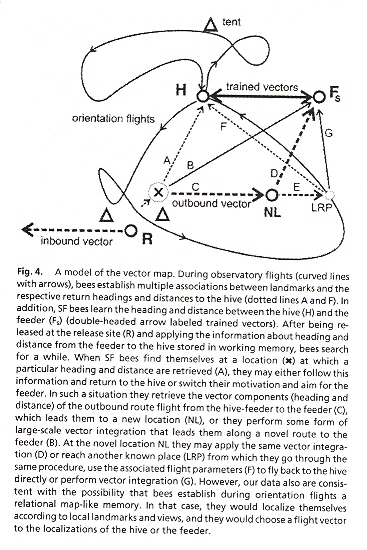
"Honey bees navigate according to a map-like spatial memory", Proceedings of the National Academy of Sciences U.S., Feb. 22 2005, 102, 8, 3040 - 3045
Two views exist about Honey bee navigation:
- Bees do NOT compute locations
- Bees have a cognitive map and do compute locations
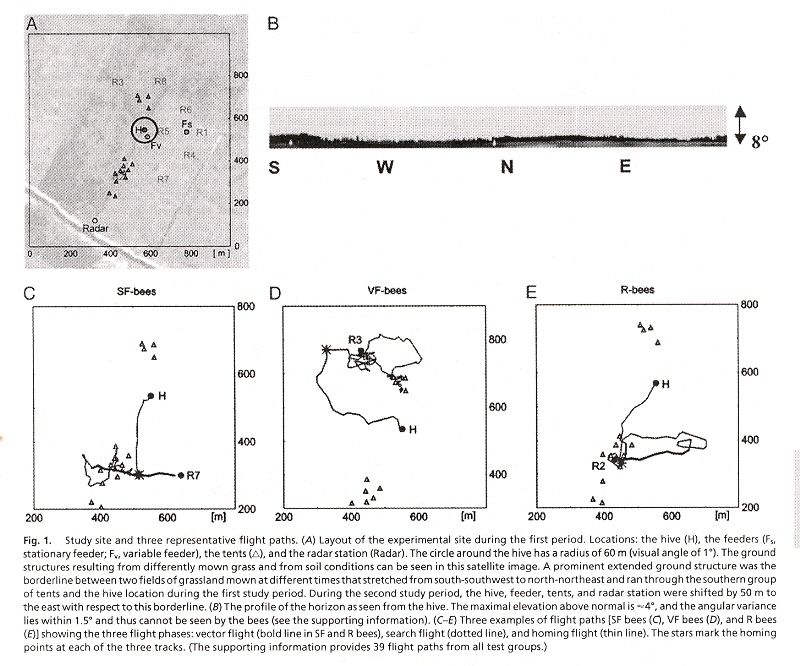
When Honey bees do not perform orientation flights (used by young bees before they start foraging), bees use a special spatial memory. Using this special spatial memory, test bees follow novel shortcuts, choosing between a feeder or the hive: a flexible form of spatial memory.
Honey bees were fitted with transponder antenna and tracked by a harmonic radar system. Test bees forage at a feeder or are recruited by waggle dance in which the location of the feeder is communicated. Flights are recorded after the bees are released at unexpected release sites. Behaviourial routines quickly emerge:
- Initial straight flight using the course when captured (foraging) or communicated during waggle dance (recruited)
- Slow search flights (to gain bearings) with frequent changes of direction
- Straight, rapid homing flights to either hive or feeding station, then hive
Bioluminescent/Chemoluminesent/Bioelectric/Biomagnetic Communication
Bioluminescence is created by either of two mechanisms: chemical bioluminescence (for example, in terrestrial Photorhabdus bacteriaa) or fluorescence. The energy to create luminescence may be generated by a chemical reactions. Fluorescence occurs when a living thing or an object absorbs light at one level of energy and then re-emits light at a lower level of energy (it is not generated). Bioluminescence is a commonly found method used for communication for a variety of purposes such as attracting mates, attracting prey, etc. Bioluminescence as a means of communication might possibly be found to have enough structure to constitute a language or to evolve to such a level of complexity. Bioluminescence occurs more frequently than one might assume. Examples include: fluorescent fungus Cnidosporidea b; fluorescent bacterium C. jejuni c; fluorescent jellyfish Aequoria victoria d; bamboo coral e; coral reef crab Lophozozymus pictor f; sea anemone Entacmaea quadricolor g; chain dogfish sharks; mantis shrimp Lysiosquillina glabriuscula h; squids i; fireflies Coleoptera: Lympyridae j; click beetles Coleoptera: Elateridae k; etc.
a "Comparison of the Bioluminescence of Photorhabdus Species and Subspecies Type Strains", by P. Hyršl, M. Čiž, A. Lojek, Folia Microbiology, 2004, 49, 5, 539 - 542
b "Rapid, hot chromotrope stain for detecting microsporidia", by O. G. W. Berlin, L. R. Ash, C. N. Conteas, J. B. Peter, Clinical Infectious Diseases, July 1999, 29, 1, 209
c "Laser scanning cytometry to detect viable but non-culturable
d "Interaction of adenosine A2A and dopamine D2-receptors as shown by FRET-technique in HEK-293 cells", by O. Kudlacek, H. Just, Naunyn-Schmiedeberg's Archives of Pharmacology, March 2002, 365, 1 (supplement), R17
e "Coral fluoresence characteristics: excitation/emission spectra, Bamboo coral fluorescence efficiencies, and contribution to apparent reflectance", by C. H. Mazel, International Society for Optical Engineering, 1997
f "Ethanolic extraction, purification, and partial characterization of a fluorescent toxin from the coral reef crab Lophozozymus pictor", by C. O. Lau, C. H. Tan, H. E. Khoo, Q. T. Li, R. Yuen, Journal of Natural Toxins, 1995, 3, 2, 87 - 90
g "The 2.0 – Aa Crystal Structure of eqFP611, a Far Red Fluorescent Protein from the Sea Anemone Entacmaea quadricolor", by J. Petersen, P. G. Wilmann, T. Beddoe, A. J. Oakley, R. J. Devenish, M. Prescott, J. Rossjohn, Journal of Biological Chemistry, 278, 45, 44626 - 44631
h "Fluorescent Enhancement of Signaling in a Mantis Shrimp", C. H. Mazel, T. W. Cronin, R. L. Caldwell, N. J. Marshall, Science, Jan. 2 2004, 303, 51
i L. Mathger, E. J. Denton, Journal of Experimental Biology, 2001, 204, 2103
j "A Field Guide to the Insects", by D. J. Borror, R. E. White, Houghton Mifflin Co., 1970, 162 - 163
k "A Field Guide to the Insects", by D. J. Borror, R. E. White, Houghton Mifflin Co., 1970, 368
"Role of molecular oxygen in the bioluminescence of the firefly squid, Watasenia scintillans, by F. I. Tsuji, Biochemical and Biophysical Research Communications, Dec. 9 2005, 338, 1, 250 - 253
Watasenia scintillans or the firefly squid is a small deep-sea squid (circa 6 cm), indigeous to northern Japan (known as hotaru-ika). These squid possess circa 800 minute dermal light organs (photophores) on their ventral sides and beneath each eye (a small number on the head, funnel, mantle and arms), but none on their dorsal sides. All the photophores emit brilliant flashes of bluish light.
"Evidence for a Category of Electroreceptors in the Lateral Line of Gymnotid Fishes", by T. H. Bullock, S. Hagiwara, K. Kusano, K, Negishi, Science, Nov. 3, 1961, 134, 1426, 1427
A description of electroreceptors is provided along the lateral line of Gymnomtus (electric eels and knife fish), and Hypopomus (electric fish).
"The Electric Sense of sharks and rays", by A. J. Kalmijn, Journal of Experimental Biology, 1971, 55, 371 - 383
Electric fields are induced by both geological and biological causes. Geological causes are based upon geomagnetic variations as well as earthquakes, etc. Biological causes emanate from living animals such as the muscular contractions of fish. These electric fields are used for navigation and environmental identification including hunting. Elasmobranchs (sharks, rays and skates) a possess electroreceptors called the "ampullae of Lorenzini". These electroreceptors have approximately the same number of nerve fibers as do those dedicated to the eyes, ears and lateral line, hence have about equal sensitivity and sensory importance to these animals. Electroreceptors in the lateral line are not as sensitive to electric fields as are the ampullae of Lorenzini, but also have an echolocation capability.
a Scyliorhinus canicula (spotted cat shark), Raia clavata (thornback ray), Galeorhinus galeus (tope shark). See "Electro-perception in Sharks and Rays", by A. J. Kalmijn, Nature, Dec. 10, 1966, 212, 5067, 1232 - 1233
Back to TopBird Communication
"The Transition to Language", Edited by A. Wray, 2002, Oxford University Press
"Sexual Displays as a Syntactical Vehicle: The Evolution of Syntax in Birdsong
and Human Language through Sexual Selection", by K. Okanoya, pages 46 - 63
It is explicitly stated in the section of this book written by Kazuo Okanoya that there is a problem reconciling the "... Darwinian process of gradual evolution and the Chomskian view of catastrophism ...". To be precise, that "... what was catastrophic was the appropriation of syntactical form for semantic purposes." Indeed, that for people such as Chomsky, "... language is functional only in its complete form." In fact, "Most studies that take a gradualist view of the evolution of language seem unnecessarily constrained from a phylogenetic perspective. For these studies, the evolution of language is the evolution of language-like behaviour in primates and hominids." Hence, an anthropomorphic constraint, as many theorists go further, to deny that language is used by anything other than the primate: the human being. The view that language may indeed apply even to non-living entities such as molecules is too radical for Okanoya. Okanoya states that the view that language is uniquely human narrows the focus severely and impedes the study of language evolution by largly excluding the examination of animal behaviour, regardless of phylogeny, that has common features with human language. Okanoya points out that both birdsong and human language share the common feature of syntax. Specifically, Bengalese finches (Lonchura striata var. domestica) sing non-deterministic songs that can be described by a finite-state syntax. Two non-deterministic FSA are provided as examples, one for the Bengalese finch, the other (far simpler), for the White-backed munia a, b, c.
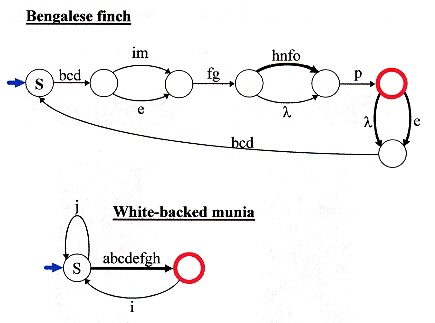
Okanoya finds both similarities as well as differences in the structure and function of linguistic centers in both bird and human brains b, and their vocal organs c (and aural organs)? Okanoya examines the development (during maturation) of syntax in both birds and humans. Okanoya concludes that early forms of hominids could have used a regular (type 3) syntax (just as the Bengalese finches display) which accounts for the evolution of hominid languages into a context-free syntax. It is the view of Okanoya that a type 3 (finite-state) syntax evolved in both the White-backed munia as well as hominids as domestication freed both from the pressure of predation to allow the sexually selective pressure to develop a linguistic (syntactic) differentiation. Okanoya then asks: "Language did not evolve in primates other than humans. Why?" Okanoya replies: "Signals take on a meaning when they can be divorced from the immediate environment of a stimulus and used to refer to some aspect of that stimulus-response event, while not actually evoking it. This requires voluntary control, and is therefore cognitive. Such behaviour emanates from the 'higher' brain, the cortex." Thus Okanoya assumes an anthropomorphic semantics that requires a brain. This is an assumption that follows when one assumes a human "conscious" state, or a cognitive state that must be implied by an anthropomorphic form of semantics, a false assumption. However, Okanoya is quite correct in pointing out that Chomsky's catastrophic viewpoint is not compatible with a viewpoint that embraces the evolution of language. One should keep in mind the dissenting views that have been explicitly stated:
- Languages and Genetics (an explicit discussion of semantics)
- The linguistic morphology found in nucleotide sequences (an explicit discussion of semantics)
- The linguistics of birdsong in courtship sexual selection (an explicit discussion of semantics)
a "Acoustical and Syntactical Comparisons between Songs of the White-backed Munia (Lonchura
striata) and Its Domesticated Strain, the Bengalese Finch (Lonchura striata var. domestica),
by E. Honda, K. Okanoya, Zoological Science, April 1999, 16, 2, 319 - 326
Six additional Finite State Automata are also provided.
b "Lesion of a higher-order song nucleus disrupts phrase level complexity in Bengalese finches",
by T. Hosino, K. Okanoya, Neuroreport, July 14 2000, 11, 10, 2091 - 2095
Six additional Stochastic Finite State Automata are also provided.
c "Song syntax changes in Bengalese finches singing in a helium atmosphere", by H. Yamada,
K. Okanoya, Neuroreport, Sept. 15 2003, 14, 13, 1725 - 1729
Five additional Finite State Automata are also provided.
"Orientation in birds", by D. R. Griffin, Experientia, 1990, 46, 335 - 336
Discussing magnetic orientation in birds, it is also noted that "...electric orientation and communication in certain fishes and echolocation by bats and odontocaete cetaceans" does exist.
"Sensory basis of bird orientation", by P. Semm, R. C. Beason, Experientia, 1990, 46, 372 - 378
Discussing the use of the Earth's magnetic field for orientation, it is noted that magnetic receptors exist for few species other than birds, such as magnetic bacteria and "... sharks and rays, which may use their electroreceptors for magnetic detection."
"Communicating about danger: urgency alarm calling in a bird", by A. J. Leavesley, R. D. Magrath, Animal Behaviour, Aug. 2005, 70, Part 2, 365 - 373
Aerial trill alarm calls of the white-browed scrubwren (Sericornis frontalis) varied according to the distance from a suddenly appearing predator: the closer the predator, the greater the number of elements in the call and the higher their minimum pitch. The experiments were confirmed with playback showing that multi-element alarm calls prompted a more urgent response. Great attention was placed upon the experimental methodology to ensure a correct interpretation of the findings. Although the bird calls were not studied from a linguistic viewpoint, the white-browed scrubwren is yet another finding that supports the view that animals other than humans have an ability to communicate meanings.
"Mechanisms of Magnetic Orientation in Birds", by R. C. Beason, Integrative and Comparative Biology", June, 2005, 45, 3, 565 - 573
Behaviour and electrophysical studies demonstrate that birds use the Geomagnetic field for migratory navigation for distances up to thousands of kilometers. Avian capabilities include direction finding (compass) as well as position finding (map). The compass capability distinguishes poleward from equatorward (rather than north from south). The receptor is located in the retina. A second receptor is based upon magnetite and is sensitive to very small magnetic field changes. It has been demonstrated that homing pigeons use a time-compensated sun-compass (compensates for the rotation of the earth), while nocturnally migrating birds use a star-pattern based compass. These capabilities are not limited to homing pigeons, but are found in other birds such as the European Robin, Bobolink, Australian Silvereyes, Pied Flycatchers, Starlings, etc. A similar ability has also been demonstrated in the eastern red-spotted newt.
"Bird Alarm Calls Size Up Predators", Science, June 24, 2005, 308, 1853 - 1855
Black-capped chickadee calls convey predator information with an elaborate system of communication of both the predator type and the degree of threat.
"Allometry of Alarm Calls: Black-Capped Chickadees Encode Information About Predator Size", C. N. Templeton, E. Greene, K. Davis, Science, June 24, 2005, 308, 1934 - 1937
An elaboration of "Bird Alarm Calls Size Up Predators".
"Dee for Danger: Chickadees add notes as threat grows", S. Milus, Science News, June 25, 2005, 403-404
Black-capped chickadee calls convey predator information with an elaborate system of communication of both the predator type and the degree of threat.
"Coevolution of Birdsong Grammar without Imitation", by K. Sasahara, T. Ikegami, 482 - 490, in Advances in Artificial Life - Proceedings of the 7th European Conference on Artificial Life (ECAL03), Edited by W. Banzhaf, T. Christaller, P. Dittich, J. T. Kim, J. Ziegler, Sept. 14 - 17 2003, 2801, Dortmund, Germany.
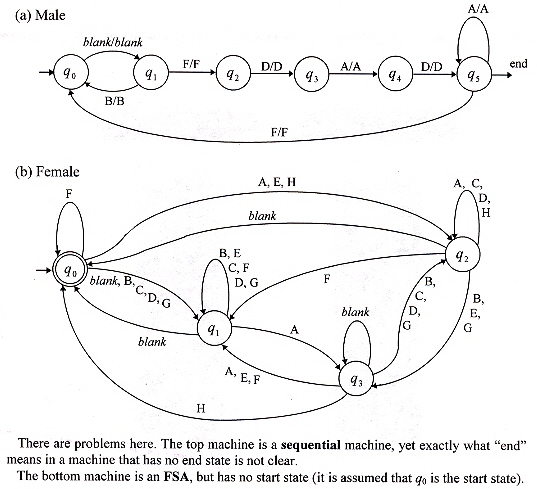
The authors point out that the song of songbirds and whales have a linguistic capability of generating infinite vocal expressions similar to that found in human beings. In this paper the Bengalese finch (Lonchura striata var. domestica) is examined. The authors conclude that a finite-state grammar (a Chomsky type 3 grammar) describes the linguistic courting behaviour of the Bengalese finch. Coevolution of male and female behaviour is accomplished with the male bird expressed as a Mealy sequential machine, and the female bird expressed as a Finite State Automaton. The authors then define the Mealy sequential machine for the male (a), defining the output set Δ = { blank, A, B, C, D, E, F, G, H }, but omit a definition of the input set. The female FSA (b) is defined, and it may thus be assumed that the Inout set Σ = { blank, A, B, C, D, E, F, G, H } = Δ. However, further difficulties present themselves as the sequential machine has an undefined "end" transition from state q5 (thereby state q5 becoming a final state, which is not appropriate for a sequential machine. The female's interjections are composed of a set { blank, # }. Evolution is modelled by modifying the state transition diagrams (nodes and transitions). The important point is that the Bengalese finch has a Chomsky type 3 grammar which is (of course) recursive, and in addition, the language has an explicit semantic interpretation (courting sexual selection).
Bengalese finch
"Mixed-species aggregations in birds: zenaida doves, Zenaida aurita, respond to the alarm calls of carib grackles, Quiscalus lugubris", by A. S. Griffin, R. S. Savan, K. Hausmanis, L. Lefebvre, Animal Behavior, Sept. 2005, 70, 3, 507 - 515
Territorial zenaida doves vigorously exclude intruders from their territory, but forage with and acquire novel foraging techniques from carib grackles. Zenaida doves have no vocal alarm signals of their own, but experimental results in this paper show that these doves do benefit from conspicuous alarm calls of carib grackles. Zenaida doves suppressed foraging in response to grackle predator alarm vocalizations. The authors point out that "The idea that doves may learn to behave adaptively to the alarm calls of carib grackles is supported by evidence that associative learning can mediate the acquisition of antipredator responses to novel acoustic stimuli."a As chickadees and sparrows have been shown to have linguistic capabilities, it might be possible for some bird species to evolve an ability to communicate using language with other bird species.
a "Antipredator responses to a previously neutral sound by free-living adult golden-mantled ground squirrels Spermophilus lateralis (Sciuridae)", by W. M. Shriner, Ethology, 1999, 105, 747 - 757
"Recursive syntactic pattern learning by songbirds", by T. Q. Gentner, K. M. Fenn, D. Margoliash, H. C. Nusbaum, Nature, April 27 2006, 440, 7088, 1204 - 1207
This paper claims to find evidence of a Chomsky context-free language capability in European starlings (Sturnus vulgaris). If A is understood to be "rattles" and B is understood to be "warbles", then European starlings are capable of recognizing and differentiating strings of acoustic sounds not only of the form (AB)n, but also of the form AnBn. For finite "n", both of these forms are regular expressions, and may be viewed as their equivalent Chomsky grammars (however, no grammars are explicitly displayed in this paper). The form AnBn can have an equivalent Chomsky context-free grammar, but constructing such a grammar, with its hierarchy including an element of obvious recursion is not very surprising (as other researchers have already published similar evidence of recursion in the linguistic capability of birds). However, the authors of this paper make much wider claims, such as:
- Languages are equivalent only to syntax (the authors make no claims for a semantic component). Indeed, without a semantics, what the authors report could be challenged as not constituting a language at all, let alone a Chomsky context-free "description".
- The authors claim that syntactic patterns such as A2B2 entail a recursive centre-embedding. However, the syntactic pattern A2B2 is of finite length, thus may also be viewed as a Chomsky type-3, finite-state grammar (thus the examples examined by the authors may all be Chomsky type-3, rather than Chomsky type-2 patterns). A far more important question arises here: exactly where is the evidence that these European starlings use a Chomsky grammar? To be explicit, where is the evidence that European starlings have a linguistic capability more closely fitting some theory of language other than Chomsky's? Exactly where is the evidence of brain structures associated with any language theory for that matter? For example, an Equal Matrix Grammar, or an L system, or even a Chomsky grammar, etc? . The only support given in evidence is the view that such linguistic structures "...must be related to the differential patterning of these elements by each grammar".
This paper supports the research of many scientists (some cited in this web site as well as in
"Emergent Computation: Emphasizing Bioinformatics"). The opposing views of N. Chomsky and
followers such as M. Hauser become ever more strained, especially as they generally ignore these
many findings. The unscientific views, based upon an anthropomorphic bias and a general
philosophy of "reductionism", will eventually be abandoned.
Unfortunately, this paper reports that European starlings required training. The
other papers cited in this web page did not require any training or other human intervention
that might bias results.
A little bird told me that birds prefer scientific views, rather than biased nonsense about
natural phenomena!
"Trained starlings can recognize their songs' grammar, study finds", by S. Borenstein, San Francisco Chronicle, April 27 2006, Science Section, p. A7. Report based upon research by T. Gentner, UC San Diego.
"Starlings' Listening Skills May Shed Light on Language Evolution", report on "Recursive syntactic pattern learning by songbirds", by T. Q. Gentner, K. M. Fenn, D. Margoliash, H. C. Nusbaum, Nature, April 27 2006, 440, 7088, 1204 - 1207, by C. Zimmer, in the New York Times, Science section D2, May 2 2006
In an interview, Zimmer quotes M. Hauser: "Memory is critical to language, because if you couldn't keep in mind several pieces of a sentence, you couldn't understand anything" and also "But memory's not specific to language". Thus the usual attitude that seeks to impose an anthropomorphic view upon all language: imposing a human semantics upon languages of birds and tamarins. M. Hauser uses artificial languages with male and female sounds. Thus languages have a new component: sexual differentiation! L = L(G, {sex}). Exactly how evolution is to apply to languages before sexual differentiation takes place is not exactly clear. Thus, the continuing error of imposing anthropomorphic views upon all languages. How shall memory and human semantics be understood when applied to DNA, RNA, or proteins? Exactly where shall scientists expect to find the "DNA mind", or the "RNA memory", or the male vs female sound perception of proteins (recall, these views are about all languages and language evolution in general)? Best to study language with scientists that do not harbour such prejudices! Zimmer also quotes Chomsky's views concerning Gentner's Starling linguistic capabilities: AnBn for finite n. Chomsky correctly points out that this "nesting" or subhierarchical and recursive capability may only be the ability to count ("n") A sequences, then count off the same number of differentiated B sequences (memory: a recursive capability coupled with the ability to count). Thus our Starlings may not have much of a linguistic recursive capability. Perhaps Starlings are obsessed with the mathematics of Peano's mathematical postulates instead?
Back to TopPhilosophy of Language
"Cognition: An Open Sandwich or an Open Question", N. S. Clayton, A review of the book "Do Animals Think", by C. D. L. Wynne, Science, July 16, 2004, 305, 344
A discussion concerning thinking (and language) in non-human animals.
"'Meaning-Making' in Language and Biology", by Y. Neuman, Perspectives in Biology and Medicine, Summer 2005, 48, 3, 317 - 327
Assuming that all languages are human languages, we might get a problem with a DNA language (a language inherrent to DNA, not just used descriptively or metaphorically by humans). The author of this paper appears to make this assumption. Immediately in the first paragraph of this paper, it appears that problems arise. For example:
- What are the metaphors and meanings expressed in a DNA language?
- What does it mean when we say DNA (language) "... metaphors are creatively generated rather than mechanically applied ...?"
- Is there a contradiction when we refer to DNA languages in a pre-given (fixed) world and a world in which evolution exists (isn't fixed)?
In general, scientific reasoning attempts to avoid ascribing 'meanings' to the things it studies (not the human languages used to describe these things and relationships). However, more problems arise. Specifically, the bottom paragraph of page 2. What exactly does it mean when we refer to the 'pragmatics' of a DNA language? Why is 'usage' synonymous with 'meaning in context'? In another paper a it is claimed that there is an isomorphism between cell and human languages. The author objects that this must include 'pragmatics' too. What exactly is the 'pragmatics' of a DNA language?
On page 3, second to last paragraph, it is claimed that living systems (is DNA always in a living system?) are reactive rather than transformatory. That transformational systems are sequential and linear. Yet modal temporal logics need not be sequential nor linear b.
Rather than continuing with this paper which more properly is one of philosophy or religion than about science, another problem arises at the last paragraph on page 3. It is claimed that we cannot understand the final conformation of a folding protein. Is it possible that there are a class of final conformations, either of equal or almost equal thermodynamic energies? A hierarchical structure might well require a unique final conformation. However, maybe living systems aren't hierarchical? For example, a language used to describe protein folded conformations might be Chomsky type 1 or Chomsky type 0 c.
There are other views that some people might find difficulty with such as "pathogens being vicious" (bottom paragraph on page 8).
a "Isomorphism between cell and human languages: Molecular biological, bioinformatics and linguistic implications", by S. Ji, BioSystems, 1997, 44, 17 - 39.
b Branching time temporal modal logics are not sequential. Modal logics which contain a circuit (are 'circular') are not linear. These modal logics are described in "Temporal Logic", by N. Rescher, A. Urquhart, Springer-Verlag, June 1, 1971.
c Chomsky type 1 or type 0 grammars are not hierarchical (tree structured), in general.
"The brain differentiates human and non-human grammars: Functional localization and structural connectivity", by A. D. Friederici, J. Bahlmann, S. Heim, R. I. Schubotz, A. Anwander, Proceedings of the National Academy of Science USA, Feb. 14 2006, 103, 7, 2458 - 2463
The main idea of this paper is to compare human language to non-human language (exemplified by non-human primates). Specifically, the functioning of the frontal operculum (FOP) in the human brain (asserted to be associated with Chomsky type-3 syntax) and Broca's area (BA 44 and BA 45) in the human brain (asserted to be asociated with Chomsky type-2 or hierarchical syntax). The functioning of the areas of the human brain are measured using functional MRI or fMRI. The non-human languages, "artificial" grammars, statistically neutral consonant-vowel syllables are visually presented to human subjects. Thus the languages considered exclude speech transmission, aural reception, and semantic contexts. Does anything stand out in this procedure as being methodologically questionable?
- Clearly, by excluding a semantics, these artificial languages are indeed very artificial, as by excluding semantics, they are not languages at all.
- fMRI does measure something, but it might be worth the time to clearly state what it measures. If the viewpoint is that fMRI measures brain activity, there is no problem. However, there might be questions concerning what linguistic brain activity is indeed measured. As the artificial languages in question are indeed not even languages, fMRI is being used to measure non-languages. Explicitly, fMRI is being used to measure the effects of visual data sans semantics. The resultant data is then promoted into the realm of linguistic data. Thus we might question the experimental methodology here. Is this not merely another example of "reductionism"?
-
It is asserted that syntactic hierarchies and recursion are
at least some of several crucial aspects that distinguish human language.
Does this assertion not beg the question of this paper? This paper asserts
to measure the results of the evolution of the linguistic capabilities of
the brain (of humans and non-humans), by excluding languages!
However, it also inserts yet another reductionist assumption. Specifically, that Chomsky grammars are the proper tool to use, and will provide an unbiased measure of both human and non-human linguistic capabilities. Thus the assumption that using only Chomsky syntactic structures will provide an unbiased measurement of anything in human brains.
However, this is far from being an unbiased assumption.
For example, the well known language (no semantics provided in this example, but a semantics can be provided a) of L = {anbncn| n > 0} is well known to be the simplest Chomsky type-1, context-sensitive language. However, this language may be viewed as an Equal Matrix Grammar, associated with a finite state automaton, thus the language used may be viewed using a different theory of syntax and as having a much simpler structure. In addition, it has been pointed out the the Matrix language below has context-free production rules in its matrices, but the language is context sensitive b. In simple terms, the theory of language used is not neutral, but inserts a bias into the level of linguistic complexity assumed. The assumption of a Chomsky theory of language must be justified, and this is not provided in this paper c, d.
Thus even if fMRI were found to measure elements associated with language, it still remains to be proven that these correspond to any specific theory of language. As to "recursion" and "hierarchies", such assumptions that these and other anthropomorphic criteria set human beings aside as being special: these assumptions are far from being unbiased. In any case, all Chomsky language types (type-3, type-2, type-1, and type-0) display aspects of recursion and hierarchical structure. -
There is yet another very basic assumption in this paper. As the authors have
been very careful to consider non-human languages, one must consider
the non-human languages in the area of animals other than non-human primates
(such as birds, cetaceans, insects, etc), as well as languages used in plants,
molecular languages such as those for DNA, RNA, and Proteins. To exclude such
languages would certainly be very biased, and as explicitly pointed out
by researchers in these areas, such languages must include not only syntax,
but their associated semantics. After all, the authors take pain to emphasize
non-human languages, then propmptly excludes them?
Exactly how does fMRI measure the brain activity of RNA using an RNA language, for example?
As RNA languages (including pseudoknots) appears to require the complexity of a Chomsky type-1 grammar (if Chomsky's theory is used), it does appear that human languages have a complexity that is simpler than RNA. Thus by one set of criteria (recursion and hierarchy), human languages are asserted to be more complex than non-human languages. By another set of criteria (RNA type-1 that have carefully been omitted), human languages are less complex than non-human languages. In this situation, we do not indeed have a measure at all!
a "Introduction to Formal Languages", by G. E. Révész, McGraw-Hill, 1983, 50 - 51
b "Regulated Rewriting in Formal Language Theory", by J. Dassow, G. Păun, ETACS Monographs on Theoretical Computer Science Vol. 18, Springer-Verlag, New York, 1989, Chapter 1, page 26
c "On Equal Matrix Languages", by R. Siromoney, Information and Control, 1969, 14, 135 - 151
d "Channel Capacity of Equal Matrix Languages", by R. Siromoney, Information and Control, 1969, 14, 507 - 511
An Equal Matrix Grammar
